Quá trình lột xác của giáp xác (tôm, cua)
Chu kỳ lột xác của các giáp xác là một trong những đặc trưng sinh lý hấp dẫn nhất của chúng. Sự hiện diện của một màng bọc vững chắc tạo thuận lợi hiển nhiên vì nó cung cấp sự bảo vệ cho cơ thể và là bộ xương ngoài cho sự đính cơ; nhưng nó đặt ra khó khăn là ngăn cản sự tăng trưởng.
Vì vậy sự gia tăng kích thước cơ thể phải xảy ra trong một loạt các bước có liên hệ đến sự lột bỏ bộ xương ngoài cũ. Ở một thời điểm được gọi là lột xác, là một biến cố có tính chu kỳ, làm gián đoạn đời sống bình thường của con vật. Ngày nay người ta nhận biết rằng các giai đoạn khác nhau của chu kỳ lột xác là nhiều hay ít liên tục, sự hồi phục từ một sự lột xác được theo sau bởi việc tích lũy các chất dự trữ trao đổi chất và sự chuẩn bị cho lần lột xác kế tiếp.
Ngay trước và sau khi lột bỏ bộ xương ngoài cũ, nước được hấp thu vào trong con vật để làm giãn nở vỏ mới còn mềm. Sự gia tăng tiếp theo về kích thước, đôi khi được nghĩ như sự “tăng trưởng”; nhưng một cách chính xác, quá trình này chỉ là một sự giãn nở cơ học. Sự tăng trưởng thực sự là sự kiến tạo mô mới, xảy ra trong các giai đoạn sau của chu kỳ lột xác.
Sự lột xác, mặc dầu chỉ chiếm một tỉ lệ ngắn của toàn thể chu kỳ, nhưng là thời kỳ có một số nguy hiểm, và tử vong thì thường cao ở thời điểm này. Các nguồn nguy hiểm tăng 3 lần thuộc cơ học, sinh lý và sinh học.
1.Khó khăn cơ học có thể được hiểu trong việc thoát khỏi vỏ cũ, các phần ngoài nở to của càng của nhiều decapod hàm chứa một vấn đề nguy hiểm đặc biệt.
2.Các vấn đề sinh lý gia tăng từ những thay đổi đáng kể các tỉ lệ ion và nồng độ ion tổng cộng của dịch cơ thể lúc lột xác, từ sự pha loãng được tạo ra bởi hấp thu nước vào trong các tế bào, và từ những thay đổi về tính thấm của bề mặt cơ thể.
3.Cuối cùng, ngay cả nếu vượt qua được tất cả các khó khăn trên, con vật vẫn phải tránh những chú ý của địch hại có ưu thế cho tới khi vỏ mới đủ cứng một cách hiệu quả để thực hiện việc trốn thoát hay có thể đối đầu. Bất cứ người nào đã từng cố gắng giữ các con cua trong một bể kính sẽ nhận thức về khó khăn phải chịu đựng của bất cứ cá thể nào không đủ may mắn để lột xác trong sự hiện diện của các đồng loại của nó.
PHẦN 1 - CẤU TRÚC CỦA VỎ
Có nhiều kiểu cấu trúc của bộ xương ngoài được tìm thấy ở giáp xác, từ các vỏ mỏng, có thể uốn cong và không được calci hóa của branchiopoda đến giáp đầu ức (carapace) cứng và dầy của decapoda. Nói chung, vật chất hữu cơ có nhiều của vỏ là một đường đa có chứa nitơ – chitin, nhưng vì chất này tương đối mềm và có thể uốn cong, vỏ được làm cứng bởi sự tẩm các muối calci và/hay bởi tanning (sự nối với nhau của các protein bằng sự thành lập các liên kết ngang giữa các phân tử kế nhau lại).
Ở decapoda (tôm, cua nói chung), vỏ lại được phân chia nhỏ thành 5 miền chính: lớp mô sừng ngoài, lớp sắc tố hóa, lớp calci hóa, lớp màng và lớp tế bào biểu mô dưới vỏ.
Lớp mô sừng ngoài (epicuticle, lớp vỏ ngoài)
Lớp mô sừng ngoài là một lớp đồng nhất nhiều hay ít, gồm protein và vật chất lipid, protein được nối bởi các liên kết ngang quinone. Lớp mô sừng ngoài có thể đáp ứng đáng kể cho việc hạn chế tính thấm của vỏ. Không tìm thấy chitin trong lớp mô sừng ngoài, nhưng lớp này đôi khi được calci hóa.
Lớp sắc tố (pigmented layer, lớp vỏ giữa)
Bên dưới của lớp mô sừng ngoài là lớp chitin được calci hóa, tuy nhiên nó cũng chứa protein liên kết ở miền phía ngoài của nó. Nhìn từ bề mặt qua lớp mô sừng ngoài, lớp này cho thấy một loạt các hình 6 cạnh. Các miền giữa của lớp chứa các trụ chitin được bão hòa với các muối calcium trong khi các protein liên kết được tập trung ở các miền gian trụ. Các bìa của các trụ có thể nhìn thấy rõ các bờ của những tế bào dưới vỏ có thể đáp ứng cho sự tiết các lớp. Các ống có lỗ có chứa các sợi nguyên sinh chất sắp xếp thẳng góc xuyên qua các trụ nhưng không có ở các miền gian trụ. Các hạt sắc tố hiện diện trong lớp này như tên gọi.
Lớp calci (calcirous layer, lớp vỏ trong)
Một lớp chitin không liên kết, bão hòa nhiều hay ít với các muối calcium nằm dưới lớp sắc tố. Nó tạo thành phần dầy nhất của bộ xương ngoài.
Lớp màng (membranous layer, lớp không được calci hóa)
Ðây là một màng chitin không liên kết, không calci hóa nằm ngay trên các tế bào dưới vỏ.
Sự phát triển tương đối của các lớp vỏ khác nhau này không ổn định ngay cả trong một cá thể. Trong các miền mềm, chẳng hạn như các miền gian đốt, vỏ mỏng hơn nơi khác, mức độ calci hóa thì nhỏ và ít có bằng chứng của sự liên kết. Trái lại, nơi mà bộ xương ngoài rất vững chắc thì không chỉ calci hóa phổ biến hơn mà còn liên kết nhiều hơn. Các vỏ của tôm cần nhẹ nhưng vững chắc có chứa mức độ đáng kể của liên kết.
5.Lớp tế bào biểu mô dưới vỏ (epidermis)
Ðây là một thành phần sống duy nhất có nhiệm vụ tiết ra các lớp trên.
PHẦN 2 - CÁC GIAI ĐOẠN CỦA CHU KỲ LỘT XÁC
Việc đầu tiên đối với sự lột xác là sự thành lập đặc biệt một lớp vỏ mới dưới bộ xương ngoài đang hiện diện và rồi các lớp giữa của bộ xương ngoài được tái hấp thu phần lớn, phần còn lại thì được lột bỏ lúc lột xác, và một sự gia tăng nhanh chóng kích thước cơ thể xảy ra trước khi vỏ mới cứng lại.
Các giai đoạn khác nhau của chu kỳ lột xác ở cua Cancer pagurus được mô tả đầy đủ lần đầu tiên bởi Drach (1939); tổng quát, có thể được chấp nhận cho tất cả Brachyura. Sự đặc trưng của các giai đoạn khác nhau như sau:
Giai đoạn A: ngay sau lột xác, không ăn
- A1: bộ xương ngoài quá mềm đến nỗi con vật không thể tự chống đỡ trên các chân của nó. Trọng lượng đang gia tăng vì nước vẫn đang được hấp thu.
- A2: sự khoáng hóa của vỏ đã bắt đầu và bây giờ con vật có thể đứng nhưng bộ xương ngoài vẫn còn mềm. Trọng lượng ổn định. Hàm lượng nước của toàn thể con vật là 86%.
Giai đoạn B: thời kỳ chính của sự khoáng hóa của vỏ mới, không ăn
- B1: sự tiết lớp đá vôi bắt đầu. Ðốt IV (đốt đùi: meropodite) và đốt trước bàn (propodite) của các chân có thể được uốn cong mà không gãy. Hàm lượng nước 85%.
- B2: sự tiết của bộ xương ngoài tiếp tục. Meropodite và propodite bây giờ nứt nếu uốn cong. Hàm lượng nước 83%.
Giai đoạn C: dầu vỏ cứng nhưng sự calci hóa vẫn tiếp tục trong những giai đoạn phụ sớm, việc ăn được tiếp tục lại.
- C1: thời kỳ chính của sự tăng trưởng mô. Các mặt đối diện của chân hơi đàn hồi khi bị nén. Hàm lượng nước 80%.
- C2: sự tăng trưởng mô tiếp tục. Các vỏ chân đàn hồi dưới áp lực nhẹ nhưng nứt nếu nén mạnh. Hàm lượng nước 76%.
- C3: vỏ cứng nhưng sự calci hóa vẫn chưa hoàn tất ở phần bên và trước của mai (carapace). Hàm lượng nước 61%.
- C4: giai đoạn “gian lột xác”. Sự calci hóa hoàn tất và lớp màng được đặt ngay phía dưới vùng được calci hóa. Các chất dự trữ trao đổi chất đang tích lũy. Sự tăng trưởng mô hoàn tất. Hàm lượng nước 61%.
Giai đoạn D: giai đoạn chuẩn bị cho lột xác tiếp theo. Sự tái hấp thu calci xảy ra và các lớp ngoài của một vỏ mới được tiết. Việc ăn ngừng lại và các dự trữ trao đổi chất được huy động. Hoạt động giảm sau giai đoạn phụ D2. Hàm lượng nước 59-61%.
- D1: dấu hiệu đầu tiên của việc báo hiệu lột xác là sự xuất hiện của những gai (setae) mới ở gốc của những gai cũ. Lớp mô sừng ngoài được tiết bởi các tế bào biểu mô dưới vỏ (hypodermis).
- D2: lớp sắc tố hóa mới được tiết.
- D3: sự tái hấp thu calci đều khắp của vỏ cũ xảy ra và điều này có thể gây ra sự nứt ở các nơi.
- D4: sự tái hấp thu calci dọc theo các đường nứt hoàn tất, kết quả tạo ra một khe của vỏ mà từ đó con vật có thể thoát ra. Hấp thu nước bắt đầu.
Giai đoạn E: lột xác. Con vật thoát ra khỏi bộ xương ngoài cũ và hấp thu nước nhanh chóng.
Tỉ lệ thời gian của mỗi giai đoạn của chu kỳ lột xác trung gian khác nhau tùy theo mỗi loài, nhưng có thể xấp xỉ: giai đoạn A = 2%, giai đoạn B = 8%, giai đoạn C = 71%, và giai đoạn D = 19%.
Mặc dầu sự phân chia chu kỳ lột xác này được thực hiện nguyên thủy chỉ trên brachyurans trưởng thành, nó cũng có thể có giá trị khi được bổ sung một cách thích hợp về chi tiết cho tất cả decapods với bộ xương ngoài dầy, được khoáng hóa tốt. Nó có thể được biến đổi để phù hợp với những cấu trúc bộ xương ngoài và sự lột xác của natantia.
Dùng tôm Caridean, Palaemon serratus, như một tiêu biểu
Thời gian của mỗi giai đoạn như sau: giai đoạn A = 2,5%, giai đoạn B = 16,5%, giai đoạn C = 21%, giai đoạn D = 60% và giai đoạn E <0,5%.
+ Ðộ dài của chu kỳ thay đổi với tuổi của con vật, loài và thời gian trong năm, và cũng có thể bị ảnh hưởng bởi nhiệt độ và tình trạng dinh dưỡng.
- Các con vật trẻ có khuynh hướng lột xác thường xuyên hơn những con già hơn.
- Sự đói thường ức chế lột xác (nhưng ở một số loài tôm nhỏ có ảnh hưởng ngược lại như tôm Leander).
- Những khác nhau về giới tính trong độ dài chu kỳ đã được chú ý ở Leander. Giai đoạn C4 chiếm một tỉ lệ của toàn thể chu kỳ nhỏ hơn và giai đoạn D có một độ dài tuyệt đối ngắn hơn ở con đực hơn là con cái. Hậu quả là con đực lột xác thường xuyên hơn con cái.
- Khi chu kỳ toàn thể rút ngắn đi, một cách tự nhiên, các giai đoạn riêng biệt cũng bị nén lại. Các quá trình bình thường xảy ra hàng ngày hay hàng giờ ở cua có thể hoàn thiện rất nhanh ở các hình thức nhỏ hơn.
- Sự gia tăng kích thước ở những con vật còn non có thể là đáng kể; nhưng cùng với sự gia tăng tuổi có khuynh hướng làm giảm sự giãn nở một cách tương ứng ở mỗi lần lột xác.
Ở các decapods có 2 kiểu chu kỳ lột xác khác nhau: lột xác theo mùa (anecdysis) và lột xác quanh năm (diecdysis). Anecdysis là sự lột xác trên cơ bản mùa hay năm. Trong chu kỳ của các hình thức này có một giai đoạn C4 dài như đã nói ở trên. Trái lại, ở diecdysis, không có sự kéo dài giai đoạn gian lột xác (intermoult). Một chu kỳ lột xác đang tham gia vào trong chu kỳ kế tiếp.
Diecdysis là đặc trưng của các loài tôm nhỏ, như Leander, lột xác quanh năm; nhưng một số cua có khuynh hướng có một số lần lột xác kiểu diecdysis trong mùa hè, được theo sau bởi chu kỳ anecdysis với một intermoult dài hơn suốt mùa đông. Một số cua sau khi đạt tới kích thước cực đại của chúng và rồi ngừng lột xác. Ðiều này cũng được gọi là “anecdysis cuối cùng” (terminal anecdysis). Sự lột xác cuối cùng không xảy ra ở tôm.
PHẦN 3 - SỰ PHÁT TRIỂN CỦA VỎ MỚI
Trước khi xảy ra sự thành lập bất kỳ một vỏ mới, vỏ cũ phải được tách khỏi phần dưới vỏ. Rồi các tế bào biểu mô dưới vỏ gia tăng về số lượng, kéo dài ra và tiết các lớp mô sừng ngoài và sắc tố của vỏ mới.
Ở cua Carcinus và tôm hùm Panulirus, lớp mô sừng ngoài được liên kết bởi quinone ngay sau khi sự thành lập của nó. Vì vậy, lớp lipoprotein đã được liên kết phân chia các lớp chitin của vỏ cũ khỏi lớp sắc tố mới có chứa chitin. Sự phân chia này có vai trò quan trọng đáng kể trong việc bảo vệ chitin mới được thành lập khỏi sự tái hấp thu vật chất từ vỏ cũ hiện đang xảy ra.
Sự tiêu hóa vỏ cũ liên tục mãi tới khi tất cả lớp màng và phần lớn lớp calci đã bị tiêu hủy. Không gian được tạo thành cho phép tách dễ dàng các vỏ cũ và mới; điều này cần thiết để con vật tự thoát khỏi vỏ cũ.
Sự tái hấp thu gần như hoàn toàn dọc theo các đường lột xác, nhưng cũng đáng kể ở gốc của các càng có phần ngoài lớn hơn. Việc tái hấp hoàn toàn phần gốc là một nhu cầu hiển nhiên để các mô của miền xa có thể thoát ra qua phần rãnh hẹp ở gốc của phụ bộ.
Ở thời điểm lột xác, lớp mô sừng ngoài của Panulirus đã hoàn thiện và lớp sắc tố đã được tiết nhưng không được liên kết vì thế nó giữ lại một mức độ đáng kể của tính co giãn.
Lượng nước cơ thể bắt đầu tăng lên rất ngắn trước khi con vật thoát khỏi vỏ cũ. Lúc này con vật trải qua một thay đổi về chất của sự co giãn và những chuyển động về phía trước khi nó nỗ lực để tự thoát. Cuối cùng sau khi chọn một vùng yên tĩnh và riêng biệt, nó uốn cong phần bụng và nhú ra xuyên qua màng gian đốt phân chia ngực và bụng.
Cơ thể được nở ra nhanh chóng trong và sau lột xác. Một đặc trưng quan trọng trong việc tạo ra khả năng nâng vỏ đầu ngực trước khi thoát ra là một sự phình ra của dạ dày. Ở một số loài điều này được thực hiện bằng cách uống nước từ môi trường nhưng ở Panulirus thực hiện bằng cách thay thế bởi những chất khí xuất hiện giữa đường nối dạ dày cũ và mới.
Phần lớn giáp xác lột bỏ toàn thể bộ xương ngoài cũ cùng một lúc. Sự lột xác ở nhiều loài xảy ra trong những giờ tối như vậy sẽ cung cấp một mức độ bảo vệ chống lại địch hại trong khi con vật không có sự trợ giúp.
Sự hấp thu nước bởi máu và các mô bắt đầu chỉ trước khi con vật thoát khỏi vỏ cũ. Khi nước đi vào cơ thể, một số được hấp thu vào tế bào và phần còn lại phục vụ để gia tăng thể tích máu. Nước hấp thu thực sự để làm giãn nở vỏ mới và các bờ không đều của các cột vật chất có đá vôi trong lớp sắc tố để cho các dạng lục giác cạnh thẳng trên bề mặt.
A. CHU KỲ LỘT XÁC TRUNG GIAN (INTERMOULT CYCLE)
SỰ KHỞI ĐẦU CỦA LỘT XÁC
Sự khởi đầu lột xác, sự bắt đầu của tiền lột xác hay giai đoạn D, ở các giáp xác decapod là dưới sự kiểm soát của hệ thống thần kinh trung ương (CNS). Sự kiểm soát này được thực hiện bởi phức hợp thần kinh-thể dịch và hệ thống nội tiết của con vật.
1. Các điều kiện bên ngoài
a. Ánh sáng
Như ở các sinh vật khác, cả hai cường độ và thời gian chiếu sáng ảnh hưởng đến giáp xác. Nếu giai đoạn tiền lột xác được khởi đầu trong một thời gian tối ổn định, ánh sáng có thể ngừng quá trình.
Một số giáp xác, hầu như sự lột xác không xảy ra trong sự tối ổn định; những thời kỳ chiếu sáng ngày dài hơn thời gian bình thường sẽ gia tăng hoạt động lột xác.
b. Nhiệt độ
Không giống như ánh sáng hay những biến đổi môi trường khác. Nhiệt độ có thể có cả hai ảnh hưởng trực tiếp và gián tiếp lên chính sự lột xác và các quá trình kiểm soát lột xác. Vì phần lớn quá trình trao đổi chất tỉ lệ thuận với nhiệt độ và hoạt động lột xác của giáp xác cũng thường gia tăng với việc tăng nhiệt độ, có lẽ do cường độ trao đổi chất của con vật đã được thúc nhanh.
Sự lột xác được tạo ra bởi sự loại cuống mắt xảy ra nhanh hơn ở các nhiệt độ cao hơn. Như vậy nhiệt độ ảnh hưởng trên toàn thể sự lột xác, không những sự khởi đầu lột xác mà còn thời gian của chu kỳ lột xác một khi nó đã được khởi đầu (L.M. Passano, 1958, không xuất bản).
Như vậy sự khởi đầu lột xác bị ảnh hưởng nhiều hơn giai đoạn tiền lột xác. Một điều thú vị là sự lột xác một khi đã được khởi đầu, có thể tiếp tục ở các nhiệt độ quá cao, mà ở nhiệt độ này sẽ ức chế sự khởi đầu lột xác.
c. Các điều kiện bên ngoài khác
Rõ ràng sự lột xác bị trì hoãn khi các con cái mang trứng. Sự hiện diện của các phôi đang phát triển trên các chân bơi ngăn chặn sự khởi đầu lột xác trong một số trường hợp, và sự lột xác thường chỉ bị trì hoãn cho tới sau thải trứng.
Ðộ mặn không có ảnh hưởng trên sự khởi đầu tiền lột xác. Có lẽ độ mặn thấp hay cao ảnh hưởng đến các cơ chế kiểm soát sự thành thục mà không ảnh hưởng đến các cơ chế kiểm soát sự lột xác, nếu những cơ chế này là phân biệt.
2.Các điều kiện bên trong
Sự khởi đầu lột xác của CNS phụ thuộc qua lại trên sự thiết lập hiệu quả các dự trữ hữu cơ, có nghĩa là một gan tụy đã tích lũy đầy đủ thường là yêu cầu đầu tiên.
Sự đói ức chế lột xác trong khi việc ăn kích thích nó. Sự lột xác có thể bị trở ngại hay ngăn cản nếu đồng thời có những yêu cầu khác đặt trên việc dự trữ thức ăn. Ðiều này giải thích tại sao trong thời kỳ thành thục tuyến sinh dục, thời gian intermoult bị kéo dài.
B. TIỀN LỘT XÁC (PROECDYSIS)
Các biến cố sinh lý chính của tiền lột xác ở decapod xảy ra khi cơ quan Y, tuyến nội tiết, được giải phóng khỏi sự ức chế của cơ quan X mà bình thường kiềm hãm nó trong thời kỳ intermoult, giai đoạn C (ở cua: C4): chúng bắt đầu tổng hợp và giải phóng hormone lột xác.
Mặc dầu giai đoạn khởi đầu này (Do) không có sự thay đổi về hình thái nhưng nó là một thời kỳ của các biến cố sinh hóa như đã đề cập ở trên. Glycogen có lẽ bắt đầu tích lũy trong các tế bào biểu mô và các mô liên kết dưới biểu mô.
Các hàm lượng calcium máu gia tăng với sự huy động từ nguồn nào đó, có lẽ là gan tụy. Phosphate gan tụy có thể giảm.
Không giống với nhiều giai đoạn khác của intermoult, sự tiến triển của các biến cố sinh lý bao gồm Do có thể bị dừng lại nếu các điều kiện bên ngoài trở nên không thuận lợi hay chúng có thể bị đảo ngược, đó là điều không thể xảy ra ở giai đoạn khác nào đó. Như vậy ánh sáng và các điều kiện không bình thường kết hợp với việc giam giữ có thể phá vỡ sự tiến triển ở giai đoạn này.
Sau đó trong tiền lột xác, bắt đầu sự thành lập bộ xương ngoài mới, ghi nhận sự chấm dứt của Do và bắt đầu của D1: đây là biểu hiện đầu tiên về sự thành lập sớm các gai mới trong lòng của các gai cũ. Vẫn sau đó, khi sự tái hấp thu gia tăng và bộ xương ngoài đang được tiết, sự thay đổi bên ngoài với những điều kiện không thuận lợi có thể làm ngừng (không lâu) tiền lột xác.
Những biến đổi về bộ xương này làm thay đổi tập tính và hình dáng của decapod: nó ngừng ăn ngay và tập tính toàn thể của nó phản ánh sự tái tổ chức căn bản bên trong với vỏ, gan tụy và máu là những mô chính đang được đề cập. Sự lột xác luôn luôn theo sau.
VỎ (INTEGUMENT)
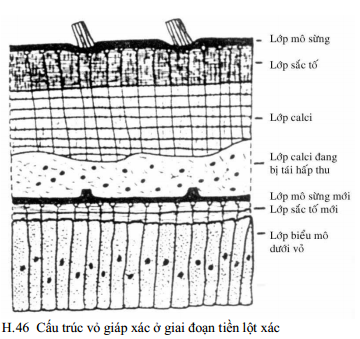
Ở cuối của sự lột xác, giai đoạn Cα (ở cua C4) vỏ bao gồm 5 lớp từ trong ra ngoài:
(1) Lớp biểu mô dưới vỏ: lớp này là thành phần sống duy nhất của vỏ và là mô có thể đáp ứng cho việc tiết các lớp còn lại.
(2) Lớp màng (lớp không được calci hóa): một lớp mỏng (20-30µ) có protein không được calci hóa, được thành lập ở cuối của C3 và nằm trên lớp biểu bì. Trong các giai đoạn tiền lột xác, nó nứt ra với sự thành lập ở gần tâm lớp dịch nhầy có tác dụng trợ giúp sự nhú ra của con vật còn mềm lúc lột xác.
(3) Lớp calci (lớp mô sừng trong): là lớp chính của vỏ. Ðây là phần chính của bộ xương ngoài, bao gồm tới 80% của độ dày toàn thể vỏ. Nó được tiết và khoáng hóa đồng thời bởi lớp biểu bì sau lột xác, trong các giai đoạn từ A2 đến C3 (ở tôm Cβ). Lớp calci có rất nhiều phiến, giàu về chitin (70% vật chất hữu cơ) nhưng nghèo về protein và thường được khoáng hóa nặng với calcium và các muối khác.
(4) Lớp sắc tố (lớp mô sừng giữa): bao gồm một số phiến chất hữu cơ cơ bản gồm 40-45% chitin và phần còn lại là protein được tạo thành trước lột xác ở D2 và D3. Sau khi lột xác, lớp này được khoáng hóa tất cả một lần bởi sự ngấm các muối. Vì lớp mô sừng giữa thường được sắc hóa mạnh mẽ, nó cùng với các tế bào sắc tố có thể đáp ứng chủ yếu cho màu sắc đặc trưng của con vật.
(5) Lớp mô sừng ngoài, một lớp không có cấu trúc về tổ chức học, cực mỏng (vài micron) được tiết bởi lớp biểu bì được hoạt hóa mới trong D1. Nó bao gồm protein được bão hòa lipid nhưng không có chitin. Chức năng của nó có thể như một lớp bảo vệ bộ xương ngoài mới đang được thành lập khỏi sự tấn công tiêu hóa của chất dịch lột xác. Nó cũng có thể tác động để hạn chế tính thấm trong các giai đoạn sau lột xác sớm.
Do là thời kỳ của sự hoạt hóa biểu bì mặc dầu không có thay đổi về mô học. Tuy nhiên, ở D1 biểu bì bắt đầu cho thấy tình trạng hoạt hóa nó một cách rõ ràng. Biểu mô tách khỏi lớp màng mà phần bên trong bắt đầu thay đổi thành lớp chứa dịch nhầy. Các dự trữ glycogen có thể chứng minh giờ đây giảm cùng lúc với sự thành lập chitin bắt đầu. Các tế bào biểu mô lớn lên, trở nên cao và phân biệt. Các dấu hiệu tổ chức học của hoạt động tiết kết hợp với sự xuất hiện của lớp mô sừng ngoài mới. Vào thời điểm D2 bắt đầu, hoạt động tiết đầy đủ được đạt tới và số lượng protein mới của vỏ đạt tới một giá trị đỉnh (E. Kuntz, 1954). Lớp giữa mô sừng được sắc tố hóa được tiết vào trong một loạt các lớp protein và chitin. Những sự đính cơ vào vỏ cũ đã đứt ra khi các cơ bị cột chặt vào bộ xương ngoài mới. Sự tái hấp thu đồng thời vật chất khoáng và hữu cơ khỏi lớp trong mô sừng được calci hóa đang xảy ra, được hỗ trợ bởi chất dịch lột xác được tiết từ biểu mô có chứa một chitinase và một phosphate kiềm. Trong giai đoạn D2 và D3, phần nhiều lớp vỏ calci cũ được loại và thành phần của nó được chuyển vào máu. Lớp giữa mô sừng (lớp sắc tố) cũ cũng có thể bị khử calci.
Sự hấp thu chuyên biệt xảy ra dọc theo đường hay rãnh trong carapace, trong các gốc chân nơi mà bộ xương ngoài nứt thành cửa lúc lột xác. Carapace bị nứt dễ dàng dọc theo đường nối này trong giai đoạn D3 nếu dùng ngón tay ấn nhẹ, trong khi nó được tái hấp thu hoàn toàn ở giai đoạn D4. Sự tái hấp thu tương tự xảy ra dọc theo các đường nối của các phụ bộ gốc chân bò cho phép việc nứt thật sự của các càng và chân bò.
Số lượng tổng cộng của sự tái hấp thu thay đổi lớn lao, không chỉ trong các decapod khác nhau mà còn trong từng loài riêng biệt với sự kéo dài tiền lột xác khác nhau. Sự tái hấp thu lớn hơn có lẽ xảy ra ở các tôm nước ngọt nơi mà calci phải được bảo vệ.
GAN TỤY
1.Các dự trữ hữu cơ
Sự tích lũy có tính chu kỳ các dự trữ hữu cơ là một đặc trưng có ý nghĩa và quan trọng của sinh lý giáp xác. Những dự trữ này, mà phần lớn chứa trong gan tụy, được dùng để thỏa mãn những nhu cầu đặc biệt về các nguyên liệu và năng lượng xảy ra trong sự lột xác, chúng không được sử dụng cho việc tăng trưởng các mô nói chung.
Sự tích lũy về các dự trữ hữu cơ đặc trưng ở giai đoạn C, đặc biệt là C4, nó tiếp tục trong Do và D1, chấm dứt khi con vật tiền lột xác ngừng ăn ở D2.
Mặc dầu gan tụy là cơ quan chính cho việc dự trữ, sự dự trữ một số glycogen cũng xảy ra ở trong biểu mô và mô liên kết dưới biểu mô. Máu cũng có thể tác động, với một nghĩa hẹp, như một cơ quan dự trữ. Các cơ cũng có thể là những vị trí dự trữ (có số lượng nhiều); chẳng hạn các hàm lượng glycogen trong các cua “cứng” giảm từ 0,2–0,6 g/100 g tới ít hơn 0,05 g/100 g sau lột xác.
a.Các lipid
Phần chính của các dự trữ hữu cơ trong gan tụy được tích lũy trong C4 bao gồm lipid, hầu như hoàn toàn ở dạng các acid béo và glycerol.
Hàm lượng acid béo của gan tụy cua Cancer được tăng 3 lần trong thời kỳ ăn mồi (giai đoạn C1 đến D1) với phần lớn tích lũy xảy ra trong C4.
Lipid hiện diện như những hạt trong “tế bào dự trữ” của gan tụy trong C4 và cũng có thể cho thấy trong các tế bào tiết (của gan tụy) trong D1 và D2. Vì cua Cancer không ăn nữa trong giai đoạn D2, sự hiện diện của các lipid trong các tế bào tiết cho thấy rằng chúng là vị trí của việc sử dụng acid béo.
Nhiều dự trữ lipid được biến đổi thành glycogen rồi thành glucose được sử dụng trong việc tạo thành chitin.
Hàm lượng cholesterol của toàn thể con vật gia tăng lúc lột xác, từ 0,036 tới 0,75 g/100 g trọng lượng ướt của mô. Sự gia tăng cholesterol tương ứng từ một giá trị tối thiểu ở C4 tới một giá trị cao sau lột xác ở cả hai: toàn thể con vật nói chung và gan tụy nói riêng.
Sự gia tăng hệ quả về hệ số lipid (tỉ lệ số lượng cholesterol/số lượng các acid béo được phosphoryl hóa) có thể gây ra đặc trưng: gia tăng sự hấp thu nước ở giai đoạn lột xác và sau lột xác sớm.
b.Các chất đường
Trong khi vai trò của glycogen trong trao đổi chất trung gian của decapod vẫn chưa rõ hoàn toàn. Không nghi ngờ rằng nó là một chất trung gian trong sự tạo thành chitin và cũng có thể có chức năng như một nguồn năng lượng cho ra sự tổng hợp vỏ.
Ở cua Cancer những số lượng nhỏ glycogen xuất hiện như những hạt nhỏ trong các vùng đỉnh của cả hai loại tế bào dự trữ và tiết của cơ quan gan tụy trong C4. Các số lượng glycogen tiếp tục gia tăng trong các phần sớm của giai đoạn D đồng thời với sự gia tăng glycogen trong biểu mô vỏ.
Glycogen biểu mô bắt đầu giảm khi sự tiết chitin bắt đầu tăng nửa sau của D1, nó tiếp tục giảm trong D2 và D3. Rồi glycogen tổng cộng của gan tụy giảm khi sự tiết vỏ gây ra sự suy kiệt nhanh hơn sự biến đổi thay thế từ các dự trữ. Glycogen gan tụy bắt đầu giảm dần dần, cùng với sự giảm hàm lượng của các dự trữ lipid; trong các giai đoạn sau lột xác, đạt tới giá trị tối thiểu của nó vào cuối của sự nhịn ăn để lột xác ở C1.
Glycogen của mô được biến đổi thành glucose mà trong quá trình thành lập chitin, nó được biến đổi thành glucosamine và acetylglucosamine. Rồi acetylglucosamine được đa hợp hóa để tạo thành chitin.
c. Các protein
Mặc dầu hiểu biết về dự trữ protein và sử dụng nó thì sơ sài, protein dường như là một thành phần cần thiết của các dự trữ hữu cơ vì nó được tích lũy trong gan tụy trong intermoult. Sau khi cho phép sự tăng trưởng mô bình thường của các giai đoạn sau lột xác (chủ yếu C1 và C2), gần 70% nitơ protein của cua Cancer tìm thấy ở giai đoạn D đã được tính toán là được sử dụng cho sự lột xác thành công. Nitơ sẽ được dùng để tổng hợp nên protein vỏ.
2. Các dự trữ vô cơ
Gan tụy có hai chức năng trong trao đổi chất khoáng: (1) nó phục vụ như một cơ quan tích lũy các dự trữ vô cơ dinh dưỡng trong C4 và (2) nó tác động như một kho tạm thời cho các vật chất vô cơ được tái hấp thu từ bộ xương ngoài cũ trong giai đoạn D.
Thành phần chính của các dự trữ khoáng ở gan tụy bao gồm các phosphate Ca và Mg. Ở cua Cancer 83,5% phosphate vô cơ là muối Ca với phần lớn còn lại là muối Mg. Các cation được sử dụng trong sự khoáng hóa vỏ sau lột xác, và phosphate thì cần thiết trong việc tổng hợp chitin trong intermoult từ C3 đến D1. Ở một số loài cua, 20-30% hay hơn nữa của trọng lượng khô của gan tụy ở D2 là vật chất vô cơ.
Calci cũng có thể được tích lũy trong máu hay nơi khác trong cơ thể. Hơn nữa có lẽ nó tích lũy trong gan tụy trong tiền lột xác trễ. Tuy nhiên, ở phần lớn decapod biển, những dự trữ lớn Ca là không cần thiết. Không giống P, Ca thì phong phú trong nước biển (khoảng 420 mg/L). Sự làm cứng sau lột xác không cần tùy thuộc vào những cung cấp bên trong, phần chủ yếu của sự calci hóa vỏ đến từ các vật chất từ môi trường bên ngoài.
Như các ion vô cơ khác, Cu cũng được giữ bởi cơ thể trong sự không ăn sau lột xác. Trong khi protein máu chứa đồng, HCy có lẽ đang được dùng như một nguồn năng lượng. Có thể các yếu tố khác chẳng hạn sulfur cũng được tích lũy.
MÁU
Chức năng chính của máu trong giai đoạn tiền lột xác là để vận chuyển các vật chất giữa vỏ và gan tụy. Nó cũng có thể tác động như một cơ quan dự trữ và như một vị trí của các phản ứng nào đó dưới sự kiểm soát của enzyme. Máu được đề cập trực tiếp với việc hấp thu nước.
Những thay đổi về các dự trữ hữu cơ ở gan tụy và vỏ gây ra những thay đổi về hàm lượng mỡ và glucose máu. Sau việc tái hấp thu vỏ bắt đầu, các hàm lượng Ca máu và P hữu cơ máu gia tăng. Protein vỏ được tái hấp thu dẫn tới một sự gia tăng protein máu.
Các hàm lượng về carotenoid, gia tăng trong máu trong giai đoạn tiền lột xác. Ở Astacus, chúng cho máu một màu phân biệt, sắc tố đó là astaxanthin.
Tốc độ đông máu thì nhanh hơn nhiều trong thời gian lột xác và máu trở nên nhớt hơn.
Trước khi lột xác có một sự gia tăng 10 lần về hàm lượng amylase máu và có một sự gia tăng về tyrosinase máu. Tất yếu rằng sự biến đổi glucose thành glucosamine xảy ra trong dịch huyết.
C. LỘT XÁC
Sự lột xác của decapod, giai đoạn E, có thể chia thành 2 phase thụ động và phase tích cực. Sự phân chia để phân biệt hai hoạt động sinh lý chính của thời kỳ này. Việc hấp thu nước được theo sau bởi sự rút nhanh chóng của con vật khỏi vỏ của nó.
Khi lột xác bắt đầu, gần như tất cả cử động của cơ thể ngừng lại, con vật đã trốn vào một nơi an toàn mà môi trường của nó cho phép. Ở phần lớn các loài, khi mà sự giao phối xảy ra trong khi con cái ở giai đoạn A1 và A2, những con cái được bảo vệ bởi những bạn kèm cứng của chúng. Ngay cả sự lột xác của natantian, tổng quát ngắn hơn nhiều so với các loài được calci hóa nặng hơn, chúng cũng ẩn mình trước khi sự lột xác bắt đầu.
Bắt đầu trong giai đoạn D4 và liên tục suốt giai đoạn E có một sự gia tăng về áp suất thẩm thấu (ASTT) của máu. Nguyên nhân của sự gia tăng này chưa rõ hoàn toàn. Ngay trước khi lột xác, có một sự gia tăng đột ngột về hàm lượng glucose máu và tiếp tục tăng trong giai đoạn A, nhưng sự gia tăng này không được tạo ra bởi một sự biến đổi chung của glycogen thành glucose cơ thể. Cả hai hàm lượng protein máu và lipid máu cũng gia tăng trước lột xác, ASTT cao hơn có lẽ được gây ra bởi những nồng độ máu tổng cộng lớn hơn của hai vật chất này.
Nhưng bất cứ nguồn gốc nào của nó, sự gia tăng ASTT máu này dẫn đến một sự gia tăng hấp thu nước đột ngột. Ở cua Cancer, và có lẽ ở các loài thủy sinh nói chung, việc hấp thu nước là ngang qua màng của ống dạ dày. Mặc dầu các tình trạng trái ngược nhau, không có sự hấp thu nước qua vỏ nói chung hay qua mang.
Sự di chuyển nước vào trong được khởi đầu khi màng dạ dày cũ vỡ ra, bộc lộ vỏ mới có tính thấm cao hơn. Nước uống vào làm gia tăng áp suất thủy tĩnh trong xoang ống tiêu hóa kết hợp với các ảnh hưởng của việc lọc và ASTT, nhanh chóng làm cho nước đi vào trong dịch huyết. Trong phase thụ động này của giai đoạn E, mà có thể kéo dài từ 2 đến 8 giờ ở Cancer, thể tích máu tăng gấp đôi. Những con vật uống nước, đẩy một phần giáp đầu ngực cũ ở đường khớp rãnh bên sau.
Khi sự rút đã đạt tới một giai đoạn nào đó, phase thụ động của lột xác được kế tiếp bởi một sự rút tích cực khỏi bộ xương ngoài cũ. Ðiều này xảy ra trong vài giây hay có thể tới 1/2 giờ, nhưng nếu bị kéo dài không bình thường, nó thường chấm dứt bằng cái chết.
Con vật không chỉ hoàn toàn không được giúp đỡ trong sự lột xác này mà còn tiêu hao nhiều năng lượng ngay ở thời điểm mà bề mặt hô hấp của nó không thể thực hiện chức năng. Tiêu hao oxygen gia tăng rõ rệt ngay trước khi lột xác và ít nhất ở cua Maja, số lượng sắc tố hô hấp trong dịch huyết đạt tới một cực đại ở thời điểm này. Dẫu sao một sự thiếu oxygen phát triển trong sự lột xác hay một biến đổi phương thức trao đổi chất kỵ khí trở nên có giá trị không được biết. Trong bất cứ trường hợp nào, khi sự lột xác được hoàn tất, con vật trở nên yên tĩnh trở lại, nhưng bơm nước qua các phòng mang của nó ở mức tối đa.
Việc hấp thu nước lại bắt đầu ngay sau lột xác, có lẽ kéo dài suốt giai đoạn A1. Nước được hấp thu xuất hiện không chỉ ở dịch huyết mà còn trong tổ chức cơ thể, đặc biệt trong gan tụy, biểu mô của màng và các cơ. Sự gia tăng hệ số lipid xảy ra ở thời điểm này, như được đề cập, để có một chức năng quan hệ nhân quả trong việc hấp thu nước này.
Sự kiện decapod lột xác tiêu biểu ở một thời điểm và một nơi mà chúng chọn riêng cho thấy sự khởi đầu lột xác của lột xác là xảy ra dưới sự kiểm soát của thần kinh. Natantians thì lột xác muộn vào ban đêm, trong khi các brachyurans như đã ghi nhận trước, thời điểm thường xuyên của sự lột xác xảy ra lúc triều thấp.
D. CÁC GIAI ĐOẠN SAU LỘT XÁC
Các giai đoạn sau lột xác của chu kỳ lột xác trung gian chiếm 70-75% hay hơn của toàn thể chu kỳ lột xác trung gian. Ở cua Maja sự calci hóa lớp vỏ giữa (lớp sắc tố) bắt đầu trong khi hấp thu nước ở giai đoạn A thì vẫn tiếp tục. Một ít giờ sau đó, những lớp đầu tiên của lớp vỏ trong (lớp calci) xuất hiện, bắt đầu một quá trình mà kéo dài đến C4.
Trong thời điểm này, thời kỳ chính của sự tăng trưởng mô cơ thể xảy ra. Cuối cùng, và đặc biệt ở brachyurans có một thời kỳ intermoult sau tăng trưởng khi cả hai: cơ thể và vỏ thì hoàn thiện và sự bổ sung của chúng đã ngừng. Ở cùng thời điểm, các dự trữ hữu cơ đang được tích trữ lại trong gan tụy trong sự chuẩn bị cho lần lột xác khác.
VỎ
Phần chính của vỏ giáp xác được thành lập sau lột xác. Lớp vỏ calci, bao gồm 75% hay hơn của bộ xương ngoài, của cua Maja thì hoàn toàn là sản phẩm sau lột xác. Sự calci hóa bắt đầu chỉ sau lột xác, sự calci hóa lớp sắc tố bắt đầu trong vòng vài giờ sau việc nhú ra của con vật còn mềm, trong khi nó vẫn ở giai đoạn A1. Sự lắng cặn calci bắt đầu với các dãy ngoài cùng nhất. Quá trình này không được hoàn thiện mãi tới giữa B1 vào thời điểm sự lắng cặn lớp vỏ calci đã tiến triển đầy đủ. Ðiều này có nghĩa rằng phần lớn sự khoáng hóa lớp sắc tố xảy ra phía ngoài đối với một lớp vỏ trong calci đã được lắng cặn và khoáng hóa. Vì thế các chất vô cơ phải đi ngang qua lớp vỏ calci để đạt đến vị trí lắng cặn của chúng. Những khoáng chất này có thể từ những dự trữ cơ thể, nhưng phần lớn được hấp thu từ thức ăn hay trực tiếp từ môi trường.
Sự lắng cặn lớp calci bắt đầu trong giai đoạn A2. Nó khác đáng kể với lớp sắc tố về thành phần, là giàu chitin và nhiều protein. Những phiến mỏng ban đầu thì dầy hơn đáng kể so với những phiến của lớp sắc tố nằm trên. Nhưng sự khác nhau đáng chú ý giữa hai lớp là rằng lớp calci được khoáng hóa khi nó được tiết. Lúc đầu sự lắng cặn nhanh, nhưng sau đó ở giai đoạn C1, chậm dần khi các dự trữ hữu cơ trong gan tụy bị suy kiệt. Trong sự tiến triển của con vật qua các giai đoạn sau lột xác, Ca có thể có giá trị từ môi trường bên ngoài.
SỰ TĂNG TRƯỞNG MÔ
Cho tới nay, chỉ các đặc trưng của sự tăng trưởng được xem xét là sự tiết của vỏ mới và sự gia tăng kích thước lúc lột xác. Ngoài những điều này, sự tăng trưởng còn liên quan đến những gia tăng về khối lượng nguyên sinh chất, được đo như sự gia tăng trọng lượng khô bắt đầu ở B2 hay C1 mà tiếp tục trong C2 và C3. Cường độ tổng hợp mới protein và DNA cao nhất trong giai đoạn này (E. Kuntz, 1954). Các mô mất nước tương ứng trực tiếp với sự gia tăng về sinh khối khô của chúng. Như vậy các mô tăng trưởng tới thể tích được thiết lập bởi sự tích nước của mô lúc lột xác.
SỰ TÍCH LŨY CÁC CHẤT DỰ TRỮ
Giai đoạn C4, giai đoạn intermoult của brachyurans và tương đương với nó trong các decapods khác, là phần của chu kỳ lột xác mà trong đó cả hai: sự thành lập bộ xương và sự tăng trưởng của mô đã được hoàn thiện. Các chất trao đổi thặng dư so với nhu cầu được chuyển đến gan tụy.
Glycogen gan tụy tăng gấp 3 và các dự trữ protein tăng, nhưng vật chất dự trữ chủ yếu là lipid. Trong toàn thể thời gian từ C1 và D1, lipid gan tụy gia tăng trên 7 lần vì thế một cua Cancer nặng 100 g ở A2 sẽ tích lũy khoảng 2 g dự trữ lipid vào lúc bắt đầu của tiền lột xác. Ngoại trừ bộ xương đã được khoáng hóa, dự trữ này khoảng 10% trọng lượng khô của toàn thể con vật. Gần như tất cả sự gia tăng này xảy ra trong C4.
Giai đoạn C4 thường được xem là “giai đoạn của sự bình thường đối với những tình trạng bị biến đổi của các giai đoạn khác”. Nó không bình thường hơn những giai đoạn còn lại của chu kỳ lột xác; và vì thức ăn thặng dư của các nhu cầu trao đổi chất đang được biến đổi thành các dự trữ được tích lũy hơn là đang được dùng hết trong sự tăng trưởng mô. Không có lý do để tin rằng một thời kỳ của tích lũy dự trữ hữu cơ là không bình thường.
Chỉ thời kỳ bình thường trong đời sống của các giáp xác là toàn thể chu kỳ lột xác trung gian như một toàn thể.
Nguồn: http://www2.hcmuaf.edu.vn